Публикации
ИНФОРМАЦИОННЫЕ ПОЛЯ И ЖИВЫЕ СИСТЕМЫ (В.В. БЕЗУГЛОВ, М.Г. АКИМОВ, С.С. КОНОВАЛОВ)
УДК 57.03 + 576.32/.36
В.В. БЕЗУГЛОВ, д.хим.н., профессор, vvbez2013@yandex.ru
Институт информационной медицины МАНЭБ, Санкт-Петербург,
Институт биоорганической химии им. акад. М.М. Шемякина и
Ю.А. Овчинникова РАН, Москва;
М.Г. АКИМОВ, к.хим.н., akimovmike@yandex.ru,
Институт биоорганической химии им. акад. М.М. Шемякина и
Ю.А. Овчинникова РАН, Москва;
С.С. КОНОВАЛОВ, д.мед.н., профессор, kvantdnk.orion777888@mail.ru
директор Института информационной медицины МАНЭБ, Санкт-Петербург
V.V. BEZUGLOV, Doctor of Chemistry, Professor, vvbez2013@yandex.ru,
Institute of Information Medicine MANEB, St. Petersburg,
Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry RAS, Moscow;
M.G. AKIMOV, Candidat of Chemical Sciences, akimovmike@yandex.ru,
Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry RAS, Moscow;
S.S. KONOVALOV, Doctor of Medicine, Professor, kvantdnk.orion777888@mail.ru
Director of Institute of Information Medicine MANEB, St. Petersburg.
ИНФОРМАЦИОННЫЕ ПОЛЯ И ЖИВЫЕ СИСТЕМЫ
Аннотация
Исходя из положения о первичности информации, постулируется наличие у любого физического объекта информационной составляющей, которую можно косвенно обнаружить в эксперименте. Цель данного исследования – проверка гипотезы, что живые системы в виде клеточных линий способны откликаться на изменения характеристики окружающих Информационных Полей (ИП), что фиксируется по изменению пролиферации клеток. Для изменения характеристики ИП используют устройство (КИП), способное фиксировать ИП в определенном объёме и тем самым увеличивать их локальную плотность. Экспозиция с КИП оказывает слабое стимулирующее действие на пролиферацию неконфлюэнтной культуры клеток. Данное действие не зависит от количества КИП, открытого и закрытого состояния КИП и времени экспозиции. Обнаруженная стимуляция сходна с действием сыворотки крови и может увеличивать выживаемость клеток в присутствии малых концентраций токсических агентов. Неконфлюэнтные клеточные культуры могут быть использованы как детекторы изменения локальной плотности ИП.
Ключевые слова: Информационные Поля, пролиферация, клеточные культуры
Summary
Based on the position of the primacy of information, it is postulated that any physical object has an informational component that can be indirectly detected in the experiment. The purpose of this study is to test the hypothesis that living systems in the form of cell lines are able to respond to changes in the characteristics of the surrounding Information Fields (IF), which is recorded by changes in cell proliferation. To change the characteristics of IF, a device (IFC) is used that is able to fix IF in a certain volume and thereby increase their local density. Exposure with IFC has a weak stimulating effect on the proliferation of non-confluent cell culture. This action does not depend on the number of IFCs, open and closed status of IFCs and exposure time. The detected stimulation is similar to the action of serum and may increase cell survival in the presence of low concentrations of toxic agents. Non-confluent cell cultures of a certain density can be used as detectors of changes in the local density of IF.
Key words: Information Fields, proliferation, cell cultures
Введение
Согласно положениям Информационно-энергетического Учения первичной является Информация. В данном случае употребляется именно такое написание этого термина, чтобы отличить его от обыденного «информация», которое относится к совокупности данных в канале передачи. Понимаемая так информация неотделима от своего носителя, будь то материя или энергия, а значит – замкнута в физическом мире.
Самое мощное предсказание в биологии
На самом деле «Информация» – значительно более глубокое и значимое понятие, позволяющее раздвинуть границы нашего познания за границы физического мира. Первым на это обратил внимание Норберт Винер, отметив, что «информация – это не материя и не энергия. Это – третья сущность». Материализм, по мнению Винера, либо должен признать этот факт, либо сойти со сцены [1]. Следует отметить, что Информация – одно из предельных понятий, определения которых возможны в частных применениях, но попытки их определить в общем виде приводят к порочному кругу замкнутых друг на друге слов.
Информация – основа и составляющая Информационного Мира (ИМ), наполняющая его Информационными Полями (ИП) различной степени сложности и качества. Физический мир возникает внутри ИМ, при его участии и остаётся под контролем ИМ. Каждый физический объект имеет информационную составляющую. Для характеристики взаимодействия ИП друг с другом не применимы понятия скорости, поскольку в ИМ нет понятия времени и расстояния. Значение имеет степень (качество) взаимодействия и завершенность или полнота результата. В случае, когда взаимодействие ИП удается частично наблюдать в нашем физическом мире, создается впечатление, что информация распространяется с чрезвычайно высокой скоростью, превышающей значение скорости света в вакууме, являющейся фундаментальной релятивистской константой. Это было убедительно продемонстрировано в эксперименте Гизина и соавт. с использованием сцепленных фотонов [2]. Сцепленные (взаимосопряженные, entangled) частицы – идентичные сущности общего происхождения и общих свойств, которые остаются в постоянном контакте друг с другом независимо от расстояния между ними. Источник, находящийся строго посередине между двумя отдалёнными детекторами, излучал пару сцепленных (t, E) фотонов в противоположных направлениях с востока на запад по оптическому кабелю. Обе ветви этой экспериментальной системы были строго одинаковой длины (17,5 км). В конце пути каждый фотон направлялся в двулучевой интерферометр Майкельсона и должен был «выбрать» длинное или короткое плечо. Независимо от числа экспериментальных тестов оба фотона «выбирали» одно и то же направление. При этом скорость информационной связи между фотонами, как минимум, в 10 000 раз превышала скорость света в вакууме.
На основе выполненных экспериментов авторы сделали однозначные выводы, что атомы могут быть соединены нелокально и даже разъединённые географически способны испытывать одни и те же изменения [2]. Таким образом, было показано, что существует возможность распространения информации иным – «нефизическим» – способом. С точки зрения ИЭУ, результаты данного эксперимента можно объяснить несколько иначе. Фотонам не требовалось обмениваться информацией и согласовывать свои действия, поскольку они фактически представляли собой материализацию одной информационной «частицы», существующей в физическом мире в нескольких копиях с одинаковым квантовым состоянием.
Чем меньше время наблюдения за событиями микромира, тем отчетливее выявляется информационная составляющая этих событий. Так, в экспериментах по измерению фотоэлектрического эффекта с помощью аттосекундной спектроскопии учёным удалось зафиксировать поведение электронов в интервале 10-19 с. При этом было замечено, что, несмотря на попытки выделить поведение отдельного электрона, результат получался такой, как если бы все электроны изучаемой системы были связаны. То есть они реагировали на экспериментальные условия как единый ансамбль [3]. Можно предположить, что при приближении к планковскому времени (5.39 x 10-44 c), которое существенно меньше достигнутой в эксперименте величины, вклад информационной составляющей возрастёт настолько, что поведение материального объекта будет практически полностью информационным.
Эти эксперименты косвенно свидетельствуют о наличии информационной составляющей материальных объектов, являющейся частью ИМ. По мере возрастания сложности структуры материального объекта возрастает сложность его информационной составляющей. У живых организмов, начиная с одноклеточных, эту информационную составляющую можно назвать Информационным Объёмом (ИО), подчеркивая сложность его организации. Благодаря особому строению ИО живых организмов, последние являются естественными детекторами информационных полей. Более того, они не могут существовать как целостные объекты без такого взаимодействия [4]. Взаимодействие живых клеток с информационными полями можно зафиксировать экспериментально.
Цель данного исследования – проверить гипотезу, что клеточные культуры могут отвечать на воздействие ИП изменением пролиферации и таким образом служить детекторами изменения свойств ИП.
Методы
Культуры клеток растили при температуре 37°С в атмосфере 5% СО2. Клетки C6, MCF-7 и HEK культивировали в среде DMEM, содержащей 10% телячьей эмбриональной сыворотки и 2 мМ глутамина. Для анализа токсичности N-ацилдофаминов культуры клеток рассеивали в 96-луночные планшеты и растили до достижения конфлюэнтности при изучении цитотоксичности и до плотности 1/2-1/10 конфлюэнтности при исследовании пролиферации. Серийные разведения испытуемых веществ в диапазоне 0.1-100 мкМ (финальная концентрация после добавления к клеткам) готовили в DMSO и растворяли в среде культивирования, после чего добавляли к культуре клеток и инкубировали 24 часа. Финальная концентрация DMSO не превышала 1%. При анализе цитотоксического действия каждую концентрацию вещества вносили в виде трех повторов. При анализе пролиферации использовали 96 лунок планшета для каждого варианта экспериментальных условий.
Для оценки пролиферации и гибели клеток под действием исследуемых веществ использовали МТТ-тест (оценка восстановления красителя МТТ митохондриями живых клеток) [4]. Для изменения характеристики ИП в локальной области пространства использовали особое устройство, названное концентратор ИП (КИП), способное фиксировать ИП в определенном объёме и тем самым увеличивать их локальную плотность. Экземпляры КИП разных размеров были изготовлены и предоставлены для данных экспериментов дмн, профессором С.С. Коноваловым. КИП способны увеличивать плотность ИП на расстоянии 5 м. По виду КИП представляет собой лист бумаги формата А4 с нанесённым типографским способом изображений и текста. Для устранения возможного эффекта опосредованного влияния компонентов типографской краски на экспериментальные образцы при проведении экспериментов каждый КИП помещали между двумя листами белой бумаги А4 и далее заворачивали в двойной лист из рекламного журнала с множеством цветных картинок и текста. В контроле вместо КИП помещали лист бумаги и упаковывали также. Далее оба экземпляра (КИП и контрольный) помещали в плотные бумажные конверты под номерами 1 и 2, которые упаковывали в пластиковые конверты с пластиковой застёжкой-молнией для обеспечения возможности стерилизации поверхности конверта перед помещение в ламинарный шкаф. Экспозицию планшетов с КИП проводили по схеме двойного слепого эксперимента. Конверты с числовыми идентификаторами были переданы сотруднику для проведения экспериментов. Вскрытие упаковки и сопоставление результатов с наличием КИП проводили в конце каждой серии экспериментов.
При изучении действия КИП при сывороточной депривации клетки растили до целевой плотности в среде с сывороткой, после этого заменяли среду на бессывороточную и проводили экспозицию с закрытым малым КИП в течение 30 мин. в CO2 инкубаторе. Оценку жизнеспособности культуры проводили через 24 часа с помощью MTT теста.
Результаты
Пролиферация клеток под действием КИП
Открытый КИП, пятикратная экспозиция
Для того чтобы проверить влияние КИП на пролиферацию клеток, была проведена серия экспериментов с серийной экспозицией с большим, открытым КИП. Экспозицию культуры крысиной глиомы С6 с плотностью 1/10 от максимальной на момент начала эксперимента проводили каждый день в течение 5 дней по 30 минут на столе, после чего культивировали клетки еще два дня и анализировали пролиферацию по сравнению с контролем. В эксперименте была зафиксирована стимуляция пролиферации на 24±3%
«Закрытый» КИП, однократная экспозиция
Для уточнения роли длительности экспозиции и открытого состояния КИП в стимуляции пролиферации была проведена дополнительная серия экспериментов с «закрытым» КИП (КИП размещался в конверте лицевой стороной противоположной той, на которой проводилась инкубация планшетов с культурой клеток) с однократной экспозицией в течение 30 мин. с последующим культивированием клеток в течение 24 ч и анализом пролиферации. Плотность клеток на момент начала эксперимента была 1/4 от конфлюэнтной. В серии зафиксирована стимуляция пролиферации на 29±7%, которая не отличается от таковой при длительной экспозиции с открытым КИП. Таким образом, однократной экспозиции с КИП достаточно для того, чтобы он оказал максимальное действие на клетки.
Стимулирующее действие КИП было проверено на дополнительной линии нетуморигенных клеток HEK 293 (Рис. 1). Экспозиция также была однократной с культивированием после неё в течение 24 ч, плотность клеток на момент начала эксперимента 1/4 конфлюэнтной. В серии зафиксировано достоверное отличие от контроля – прирост на 8±8%.
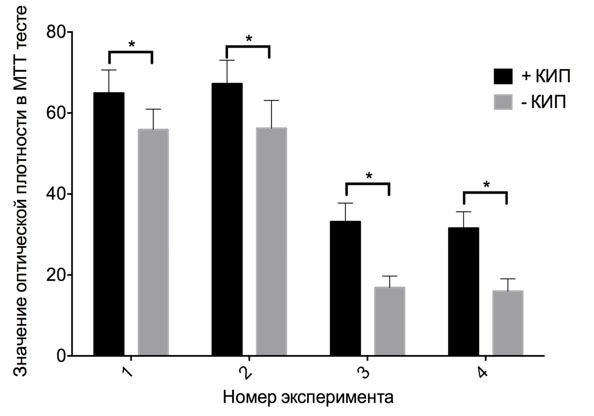
Рис. 1. Пролиферация культуры клеток HEK 293 под действием КИП. * - достоверные отличия по результатам дисперсионного анализа с пост-тестом Сидака, p < 0.05.
Влияние числа КИП
Для того чтобы проверить наличие кумулятивного действия КИП, была проведена серия экспериментов по сравнению эффекта от экспозиции одного и двух КИП. При экспозиции с двумя КИП первый был расположен под планшетом с клетками, а второй - над планшетом лицевой стороной к планшету. Время экспозиции (однократно) составляло 30 минут, анализ пролиферации проводили через 24 часа, плотность клеток была 1/4 конфлюэнтной. Достоверных отличий между воздействием одного или двух КИП не обнаружено.
Зависимость действия КИП от плотности клеток
Для того чтобы проверить, влияет ли КИП на деление клеток или на интенсивность внутриклеточных процессов, было проверено влияние экспозиции на культуру клеток глиомы с разной плотностью: 100 000, 50 000, 25 000 и 12 500 клеток/см2 на момент начала эксперимента (от конфлюэнтной до 1/8 конфлюэнтной, соответственно) при однократной 30-мин экспозиции с анализом пролиферации через 24 часа. Достоверное влияние обнаружено только при плотности клеток 50 000/см2 или менее. При этом чем меньше была плотность культуры, тем более выраженным был эффект (Рис. 2).
Для конфлюэнтной культуры влияние КИП на пролиферацию не было зафиксировано ни для одной из линий C6, HOS и HEK 293.
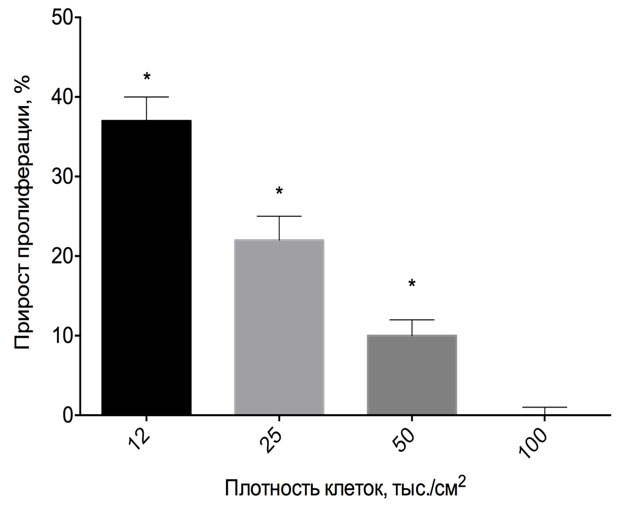
Рис. 2. Влияние КИП на пролиферацию культуры клеток HOS разной плотности. Клетки рассеивали в 96-луночный планшет и растили до плотности 32, 16, 8 и 4 тыс. на лунку. После этого экспонировали с КИП в течение 30 мин. при комнатной температуре и в обычной атмосфере, культивировали в CO2 инкубаторе 24 ч. и анализировали с помощью MTT теста. Прирост сигнала по отношению к контролю, данные репрезентативного эксперимента, среднее ± стандартное отклонение. * - достоверное отличие от контроля, дисперсионный анализ с пост-тестом Тьюки, p<0.05.
Защита от неблагоприятных условий с помощью КИП
Гибель С6 под действием ацилдофаминов
Поскольку было зафиксировано стимулирующее действие КИП на пролиферацию клеточных культур, была проверена его способность защищать клетки от действия неблагоприятных факторов - цитотоксических дофаминамидов олеиновой (Ol-DA) и арахидоновой (AA-DA) кислот [5]. Экспозицию с большим КИП проводили однократно в течение 30 мин. сразу после добавления цитотоксического агента, пролиферацию анализировали через 24 часа. Значимого влияния экспозиции в присутствии КИП на гибель клеток под действием Ol-DA обнаружено не было. Однако в случае AA-DA зафиксировано снижение гибели клеток после экспозиции на КИП на 20-30% при концентрации вещества менее 3 мкМ (Рис. 3).
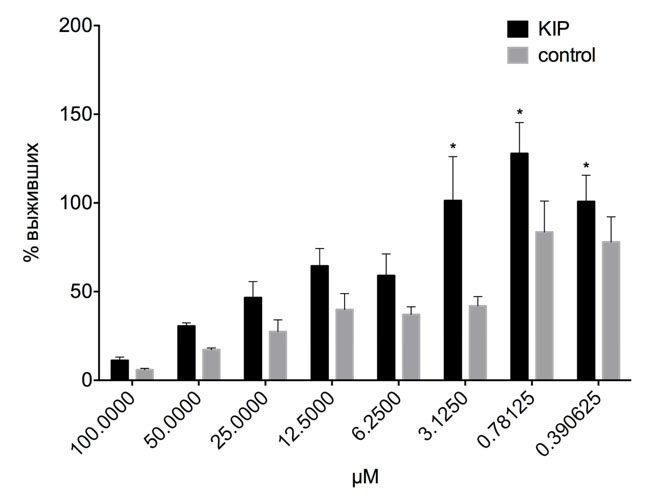
Рис. 3. Культуру клеток глиомы C6 инкубировали с обозначенными концентрациями AA-DA в течение 22 ч после экспозиции с КИП в течение 30 мин. Данные МТТ теста. * - достоверные отличия по результатам многомерного дисперсионного анализа с пост-тестом Сидака, p < 0.05.
Сывороточная депривация
Сыворотка крови является стандартным компонентом сред для культивирования клеток млекопитающих. Без неё деление клеток прекращается из-за отсутствия сигналов содержащихся в ней факторов роста, а для некоторых культур может запускаться механизм клеточной смерти [6]. Поскольку обнаружено, что КИП стимулирует пролиферацию клеток, было решено проверить, может ли он имитировать действие компонентов сыворотки. При экспозиции культуры клеток в отсутствии сыворотки, но при наличии КИП была зафиксирована стимуляция роста на 12±1% (Рис. 4). При экспозиции на КИП непосредственно среды без клеток, как в течение 30 мин., так и в течение трёх суток, статистически достоверного влияния на рост клеток в этой среде не обнаружено.
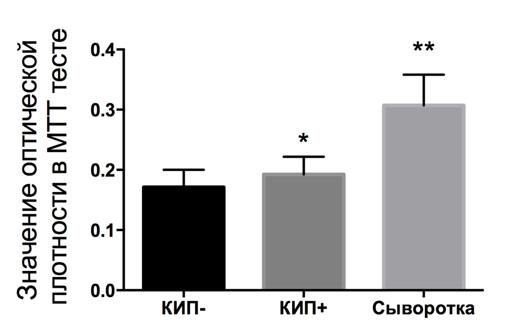
Рис. 4. Действие КИП на пролиферацию культуры клеток глиомы C6 на фоне сывороточной депривации. *, ** - статистически достоверное отличие от контроля (без сыворотки и без КИП) по результатам дисперсионного анализа с пост-тестом Тьюки, p<0.05.
Заключение
Таким образом, экспозиция с КИП оказывает слабое стимулирующее действие на пролиферацию неконфлюэнтной культуры клеток. Данное действие не зависит от количества КИП, способа позиционирования КИП (лицевой или обратной стороной) и времени экспозиции (менее 30 мин. не проверяли). Важно отметить, что максимальный прирост пролиферативной активности составил 29±7% и не зависел от длительности экспозиции культуры клеток в присутствии КИП. Можно предположить, что полученное значение может характеризовать степень снижения плотности ИП планеты, что отражается на неоптимальном функционировании клеточных систем. Введение клеток в более плотные ИП восстанавливает изначальное состояние функциональных способностей клетки, что отражается на её пролиферативной активности. Следует отметить, что хотя линии раковых клеток более активно откликались на действие ИП, увеличение пролиферативной активности было отмечено также и в случае нормальных клеточных линий эмбрионального происхождения. Обнаруженная стимуляция пролиферации в присутствии КИП сходна с действием сыворотки крови и может увеличивать выживаемость клеток в присутствии малых концентраций токсических агентов. Для реализации эффекта необходима экспозиция КИП непосредственно с клетками. Интересно, что конфлюэнтные культуры клеток были практически нечувствительны к плотности ИП. Возможно, в таком состоянии клетки уже не могут увеличивать пролиферативную активность, а другие параметры биохимии клеток в данном исследовании не измеряли. Неконфлюэнтные клеточные культуры могут быть использованы как детекторы изменения локальной плотности ИП.
ЛИТЕРАТУРА
- Wiener N. (1961). Cybernetics, or control and communication in animal and machine./ 2nd Ed. Cambridge, MA: MIT press, P. 132.
- Salart D., Baas A., Branciard C., Gisin N., Zbinden H. Testing the speed of 'spooky action at a distance'. // Nature. — 2008. — V. 454. — P. 861–864.
- Ossiander M., Siegrist F., Shirvanyan V., Pazourek R., Sommer A., Latka T., Guggenmos A., Nagele S., Feist J., Burgdörfer J., Kienberger R., Schultze M. Attosecond correlation dynamics // Nature Physics. — 2017. — V. 13 — P. 280 –286.
- Mosmann T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J Immunol Methods 1983;65(1-2):55-63.
- Akimov MG, Bezuglov VV. N-Acylated dopamines: A new life for the old dopamine. In: Kudo E, Fujii Y, editors. Dopamine: Functions, Regulation and Health Effects. NY: Nova Science Publishers; 2012. p. 49-80.
- Jakoby WB, Pastan IH. Cell culture. New York: Academic Press; 1979.
REFERENCES
- Wiener N. (1961). Cybernetics, or control and communication in animal and machine./ 2nd Ed. Cambridge, MA: MIT press, P. 132.
- Salart D., Baas A., Branciard C., Gisin N., Zbinden H. Testing the speed of 'spooky action at a distance'. // Nature. — 2008. — V. 454. — P. 861–864.
- Ossiander M., Siegrist F., Shirvanyan V., Pazourek R., Sommer A., Latka T., Guggenmos A., Nagele S., Feist J., Burgdörfer J., Kienberger R., Schultze M. Attosecond correlation dynamics // Nature Physics. — 2017. — V. 13 — P. 280 –286.
- Mosmann T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J Immunol Methods 1983;65(1-2):55-63.
- Akimov MG, Bezuglov VV. N-Acylated dopamines: A new life for the old dopamine. In: Kudo E, Fujii Y, editors. Dopamine: Functions, Regulation and Health Effects. NY: Nova Science Publishers; 2012. p. 49-80.
- Jakoby WB, Pastan IH. Cell culture. New York: Academic Press; 1979.